両性および単性の花
花は両性(アンドロエシウムと雌蕊群)または単性(雄蕊のみまたは雌蕊群のみ)です。 単性花は、オーク、シラカバ、トウワタ、トウモロコシなどの同じ植物にある場合もあります (その場合、植物全体が両性になります)。または、ポプラ、ヤナギ、麻などの別の植物にある場合もあります (その場合、雄株と雌株が存在します)。 。 これに関して、植物学文献には雌雄異株と雌雄異株という 2 つの用語が長い間存在していました。 リンネの時代以来、多くの植物学者がこれらの用語を植物自体に適用し、雌雄異株と雌雄異株の植物について語ってきました。 多くのキク科のように、植物に両性花と単性花の両方が含まれている場合、それらは一夫多妻制であると言われます(ギリシャ語のポリ(多)とガモス(結婚)に由来します)。 しかし、O. P. de Candolle、S. L. Zndlihor、D. Weptham、J. D. Hooker から A. Engler、R. Wettgaten、A. B. Repdl、J. Hutchinson まで、多くの著者が「雌雄異株」という用語を使用してきました。 植物全体ではなく、花に対してのみ「雌雄同株」です。 これら 2 つの用語の使用法のどちらがより正しいかについて時々生じる論争は、本質的には無意味です。 麻や柳について 平等基準雌雄異株であるとか、花が雌雄異株であるとか言います。 文脈によっては、これらの用語のどちらかを使用した方が便利な場合がありますが、これが誤解を招くことはありません。
単性花が両性花から生じたと信じる十分な理由があり、単性花では、雌雄異株は単複性よりも明らかに遅い。 前世紀後半以来、比較形態学と受粉生物学に関する数多くの研究により、単性花は、ある場合には雄しべ、ある場合には心皮の発育不全または完全な抑制の結果として両性花から生じたという結論に達しました。 多くの属や科全体の単性花では、雄しべと心皮(いわゆる雄しべと心皮)の縮小した残骸(原始体)が保存されることがよくあります。 このような残留形成は、プラタナス、一部の桑、イラクサ、クルミなど、さまざまな科の代表の花で見られます。 基本 生物学的原因チャールズ・ダーウィンがかつて指摘したように、両性花から単性花への移行は、より信頼性の高い他家受粉によるものです。
これらの行を読んだ後、読者は次のような質問をするかもしれません。花は胞子体の一部、つまり無性世代の一部であり、したがって性別がないのに、花の性別について語ることは可能でしょうか? 一部の植物学者はそう信じており、「男性」、「女性」、「両性」という用語の代わりに、「雄しべ」、「雌しべ」、「完全」(雄しべと心皮の両方があるという意味で完全)という用語を使用することを好みます。 )。 しかし、ほとんどの植物学者は「バイセクシュアル」と「ユニセクシュアル」、「男性」と「女性」という用語を使い続けていますが、それには十分な理由があります。 形態学的には、花は間違いなく胞子体の一部ですが、機能的には性的過程に直接関係しています。
私たちが雄花と雌花について話すとき、それは有性生殖の準備におけるそれらの役割を意味しており、有性世代 (配偶体) に属するものではありません。 要点は、男性と女性の性の間の遺伝的および生理学的分化が無性世代に伝達され、胞子の特定の有性化が起こるということです。 これは雌雄異株の植物(雌雄異株の花を持つ植物)で特に顕著です。 大麻植物の雄と雌は遺伝的および生理学的に異なり、雄の大麻はそうではないとさえ言えるかもしれません。 程度は低いが動物では雄よりも雄。 同じ理由で、雄しべは雄の構造であり、心皮は雌の構造であると考えることができます。
植物の一生: 6 巻。 — M.: 啓蒙です。 A.L.タクタジアン編集、 編集長メンバー修正 ソ連科学アカデミー教授 A.A. フェドロフ. 1974 .
サブクラス Magnoliidae サブクラス Magnoliidae サブクラス Magnoliidae には、最も古い現存する顕花植物が含まれます。 モクレン科の主要かつ中心的なグループであるモクレン科は、特に原始的なものです。 しかし、いくつかの原始性を想像してみてください。
マンサク科 (マンサク科) マンサク科 (マンサク科) マンサク科について、その中で重要な位置を占めるマンサク科から始めます。 これは古代の家族であり、その最大の繁栄の時代は第三紀でした。 私の見せ方
顕花植物の分類と系統 顕花植物の分類と系統 顕花植物を分類する最初の試みは次のとおりです。 フローラ一般に、これらは、いくつかの、恣意的に取られた、容易に認識できる外部の兆候に基づいていました。 これらは純粋に人工的なクラスでした
サブクラス DILLENIIDAS (DILLENIIDAE) サブクラス DILLENIIDAS (DILLENIIDAE) Dilleniids は、顕花植物の最大のサブクラスの 1 つです。 系統発生学的にも、最も重要な枝の 1 つです。 家系図、これはモクレン類とロシド類の間のリンクです。 の上
ロシッドサブクラス (バラ科) ロシドサブクラス (バラ科) ロシッドサブクラスに含まれる目は、外観、花の構造、栄養器官の解剖学的構造が大きく異なります。 ただし、それらは共通の起源によって結合されており、他のサブクラスと同様に、ロシドは自然な性質を表します。
サブクラス ユリ科 (ユリ科) サブクラス ユリ科 ユリ科は、すべての最大の科を含む単子葉植物の大きなサブクラスです (次のサブクラス アレシ科に属するヤシ科とアルマ科を除く)。 ユリ科の中には比較的原始的な植物があり、
ポリガミック ポリガミックは、両性花と単性花が同じ植物、または同じ種の異なる標本で見つかる現象です。 植物用語の辞書。 — キエフ: ナウコヴァ・ドゥムカ。
Sc博士の総編集の下で。 I.A. パイプ。 1984年。英雄的な 雌しべと雄しべの両方を持つ花(アンドロエシウムと雌蕊)と呼ばれます。 場合によっては、この用語は両性花にも適用されます 完璧 または 雌雄同体
花。 雄しべだけ(アンドロエシウム)または雌しべだけ(雌しべ)を持つ花を「雌雄同体」と呼びます。同性 。 雄しべのある単性花は、完璧 スタミナテ、男性用 フラワーズ; それぞれ、雌しべだけの花 -完璧 めしべ、.
雄花と雌花の単性花が同じ植物に生育することがあり、その場合、その植物はと呼ばれます。 雌雄同体、 または バイセクシュアル、 例えば: オーク、バーチ、トウダイグサ、トウモロコシ。 この場合、受粉は同じ植物内の花の間で発生する可能性があります。
雄花と雌花が異なる植物に成長する場合、私たちは次のことに対処します。 雌雄異株植物。 スタミナ状の花を持つ雌雄異株の植物を 男 そして女性と – 女性 植物、例えば: ポプラ、ヤナギ、麻、イラクサ。 雌雄異株の種の受精には、雄と雌の異なる性の植物が少なくとも 2 つ存在する必要があります。
両性花と単性花の両方を持つ植物を「植物」といいます。 一夫多妻制, たとえば、そのような近傍はキク科の花序に見られます。
胞子をもつ器官を持たない花は、 無菌、 または 無性愛者キク科の花序にある舌状花などの花。
一夫多妻植物の例:写真のガーベラの花序には、雄花(黄色い葯を持つ)、雌花(白い雌しべを持つ)、そして縁に沿って無菌の葦の花があります。
穀物とスゲの花。
穀物とスゲの花。
穀物の花は通常小さくて目立ちません。 風による受粉に適応しているため、昆虫を引き寄せる理由がないため花被がありません。 穀物の花は小穂の側枝にあり、次のもので構成されています。 雄しべ そして 卵巣 と 柱頭の枝 。 花は上下で守られている 花の鱗 。 花の鱗片の上には、2つの小さな無色の鱗片が成長します。 花の映画 、 または 嘲笑 。 開花中、長い雄しべが鱗片を越えて突き出し、風に乗って花粉を散布します。 穀物の花は両性または単性であることがあり、同じ花序内にある場合もあります。
スゲの花も小さくて目立たず、さまざまな小穂に集められ、「スゲ」と呼ばれる苞の葉腋に位置します。 スケールを覆う 。 スゲの花自体は次のもので構成されています。 雄しべ そして hネクタイ と 柱頭の枝 。 花は両性および単性であり、花被の有無にかかわらず。 スゲの花被片は、鱗片の配列、毛深いまたは縁取られた剛毛、あるいは絹のような毛で構成されており、両性花または雌花に多く存在します。
アンドロシウム
(ギリシャ語 「男の家」): 全体 小胞子葉、雄しべは 2 つの半分に分かれた糸状体からなる 葯 4つ入っている 小胞子嚢 (花粉嚢)。 雄しべは1つまたは2つの円を描くように配置されています。 雄しべは遊離雄しべと融合雄しべに分かれます。
アンドロエシウムにはさまざまな種類があり、融合した雄しべの数によって区別されます。
-兄弟らしい(雄しべが 1 つのグループにある、ルピナス、ツバキ)、
-二者関係の(2つのグループの雄しべ)、
-多兄弟(いくつかのグループ、マグノリア、セントジョーンズワート)、
-兄弟的な(融合していない雄しべ)。
雄しべの長さもさまざまです。 等しい, 不平等, 2馬力(雄しべ4本のうち2本が長い)、 3馬力(雄しべ6本のうち3本が長い)、 4馬力(雄しべ6本のうち4本が長い)。
おしべからなる 雄しべ組織、その上端に位置します。 葯、および下端がレセプタクルに取り付けられています。 フィラメントの主な組織は、 実質。 重要なプロセスは葯で発生します - 小胞子形成(小胞子嚢における小胞子の形成)および 微小配偶子形成(雄性配偶体の小胞子からの形成)。 無菌のおしべはこう呼ばれます スタミノデア.
図3 おしべと葯の発達

葯表皮に囲まれた均質な細胞で構成されています。
ダイアグラム- これは花を平面上に投影した概略図であり、花はその軸に対して垂直に横に交差しています。 図のデザインのルール: 花序の軸が上部、葉が下部で覆われます。 図の記号: 円弧は花被の一部を示し、がく片 - 円弧の中央に突起があるもの、花弁 - 突起のないものを示します。 雄しべは、葯または花糸の断面によって示されます。 雌蕊は卵巣の断面の形をしています。 個々のメンバーが一緒に成長する場合、これは図上の円弧で示されます。
アンドロエキウム
アンドロエクウムは雄しべの集合体であり、そこで小胞子形成、微小配偶体形成、雄胞子の形成が起こります。
個体発生中に、雄しべは次のように成長円錐の結節の形で形成されることがあります。 頂弁状(つまり、ベースからトップまで) 基底弁(上から下へ) シーケンス。 前者の場合、最も若い雄しべは花の中心近くに位置し、最も古い雄しべはその周辺に近く、2番目の場合はその逆です。 雄しべは、さまざまな方法で自由または融合することができます。 程度は様々ですが。 たとえば、熱帯のセンダン科では、10 本すべての雄しべが糸とともに管の中に成長します ( 兄弟らしいアンドロエキウム)。 セントジョーンズワートでは、雄しべが束になって成長し、キク科の植物は葯が接着するのが特徴です。 マメ科の多くの代表的な植物では、9 本の雄しべが一緒に成長し、1 つは自由のままです (いわゆる 二者関係のアンドロエキウム)。
各雄しべは、細くなった糸状、またはまれにリボン状または花弁状の部分である花糸と、通常は拡張した部分である葯で構成されています。 ブーツには 2 つの半分が互いに接続されています 連絡担当官、フィラメントの続きです。 靱帯は声門上まで続くこともあり、葯の上の小さな突起として目立ちます。
花糸の形成は葯よりも遅く始まり、挿入成長によりさらに伸長します。 形成される結節の数は時々 少ない数雄しべ、後に結節が分かれ、かなりの数の雄しべ(ミモザ)ができることがあります。 雄しべの花糸の長さ さまざまな植物不定。 多くの場合、それらの長さは花被とほぼ同じですが、よく知られている熱帯植物のように、花被よりもはるかに短いか、何倍も長い場合もあります。 薬用植物シソ科のキドニーティー、または猫のひげ。 フィラメントの断面を見ると、その大部分が実質組織で構成されており、中心に 1 本の維管束が走っていることがわかります。
葯の各半分には 2 つの (まれに 1 つ) 巣ができます。 小胞子嚢。 葯の巣は花粉嚢と呼ばれることもあります。 成熟した葯では、巣間の隔壁はほとんどなくなります。 葯の外側は表皮で覆われています。 表皮のすぐ下には、と呼ばれる細胞の層があります。 内皮細胞、二次的に細胞膜が厚くなっています。 内皮膜が乾燥すると、葯の巣が開きます。 1〜3層の中型の薄壁細胞がより深いところにあります。 花粉嚢の空洞を裏打ちする最も内側の層は、と呼ばれます。 タペーマ。 その細胞の内容物は、母細胞を発達させるための栄養として機能すると考えられています。 小胞子(小胞子細胞)およびその分化を促進します。 葯の巣は通常、小胞子母細胞、小胞子、成熟した花粉で満たされています。 知られているように、小胞子は減数分裂の結果として小胞子細胞から発生し、小胞子細胞自体は始胞子層(葯巣発達の初期段階で機能する教育組織)のいくつかの細胞から発生します。 熟した葯は、縦方向の亀裂、穴、弁など、さまざまな方法で開きます。この場合、花粉がこぼれます。
構造、形状、位置、雄蕊の数の兆候、およびアンドロエシウム自体の種類は、開花植物の体系化とその系統発生の知識にとって非常に重要です。
いくつかの種では、雄しべの一部が本来の機能を失い、無菌状態になり、いわゆる雄しべになります。 スタミノード。 葯は、蜜を分泌する花の分泌部分である蜜腺に変化することがあります。 花びら、その一部、雌しべの一部、さらには花托の伸長物も蜜腺やオスモフォアに変化することがあります。 蜜腺にはさまざまな形があり、通常は花の奥深くに位置し、光沢のある表面によって区別されることがよくあります。
雌蕊群。
1 つ以上の雌しべを形成する 1 つの花の心皮の集まりをと呼びます。 雌蕊園。 心皮、または 手根皮、起源は葉に関連していると考えられている構造です。 しかし、機能的および形態学的には、心皮は栄養葉ではなく、大胞子嚢、つまり大胞子葉を有する葉に対応します。 ほとんどの形態学者は、進化の過程で、平らで開いた心皮が、折り畳まれた心皮に進化したと信じています。 重複する)心皮。 その後、それらは端で一緒に成長し、最も重要な部分である胚珠を運ぶ子房を備えた雌しべを形成しました。 このようにして、どの植物群にも見られない、確実に保護された胚珠が発育する密閉容器に似た独特の構造が形成された。 雌しべの構造は受粉と受精に理想的に適合しています。 子房に位置する胚珠では、大胞子形成と巨大配偶体形成のプロセスが起こります。
雌しべ、あるいは子房は、胚珠を乾燥から守る湿った部屋として機能します。 これにより、被子植物は環境湿度のレベルに実質的に影響されなくなり、乾燥地帯の広範な開発の要因の 1 つとなった。 さらに、雌しべは胚珠を昆虫による食害や部分的には温度変動から確実に保護します。
1つの心皮から形成された雌しべを単純と呼び、2つ以上の融合した心皮から形成された雌しべを複合と呼びます。 単純な雌しべは通常、単葉です。 複合体は複数の巣に分割することも、単一の局所にすることもできます。 多房性は、心皮の融合の結果として、または追加の隔壁の形成、つまり子房の壁の成長の結果として発生します。
雌しべの柱頭は、花粉を受け取るために設計された顕花植物にのみ特有の構造です。 それは花柱の上部、または子房に直接発生します - 固着柱頭。 それほど頻繁ではありません(古種では) - 心皮の融合した端に沿って。 柱頭の形や大きさが違う 他の種類。 柱頭の表面は、多くの場合、でこぼこした塊状で、粘着性の液体で覆われており、花粉のより効果的な固定と捕捉に貢献しています。 さらに、柱頭の表面には薄いタンパク質の層である薄膜があり、花粉粒の胞子皮のタンパク質と相互作用して、花粉管の発芽を確実にしたり、発芽を妨げたりします。
この柱は緩い実質組織で構成されています。 それは柱頭を上に持ち上げるように見えますが、これは受粉プロセスのいくつかのメカニズムに必要です。 柱の形態は非常に多様であり、重要な体系的な特徴として機能します。 多くの古期の科(特にモクレン科の亜綱)は、スタイルの欠如または弱い発達によって特徴付けられます。 たとえば、多くの穀物など、多くの特殊な風受粉形式では、花柱が未発達であることがよくあります。 風によって受粉される大きな花(一部の種類のユリ)では、柱がかなりの長さに達し、柱頭が高いところに運ばれるため、受粉が促進されます。 しかし、これにより花粉管の経路が大幅に長くなります。
子房は雌しべの最も重要な部分であり、胚珠を持ちます。 形も様々ですし、 外観、これは主に雌蕊群の種類によって決まります。
胚珠が子巣に付着している場所を「卵巣」といいます。 胎盤。 胎盤は通常、卵巣の組織によって形成された小さな腫れ、成長物、または突起のように見えます。
卵巣の壁への胚珠の付着の特徴に応じて、いくつかの種類の胎盤形成が区別されます。
· 壁、または頭頂、胚珠が子房の内部の壁に沿って、または心皮が一緒に成長する場所に位置する場合。
· 軸状、または軸状、胚珠が子房の中央の柱に位置する場合、心皮の数に応じて巣に分割されます。
・自由中央胎盤、胚珠が卵巣壁に隔壁によって接続されていない自由中央柱上で発育する場合。
· 基底、唯一の胚珠が単房子房の基部に位置する場合。
婦人科の種類:
1. 無果皮 - 心皮は一緒に成長せず、各心皮が別々の雌しべを形成します (キンポウゲ、バラ)
a) 単量体 - 雌蕊は 1 つの雌しべからなり、1 つの心皮 (エンドウ豆、プラム、サクランボ) によって形成されます。
b) 高分子 - 雌しべは多数ありますが、それぞれが 1 つの心皮で構成されています。
(キンポウゲ、イチゴ、ローズヒップ)
1. 共実果 - 雌しべは心皮が融合して形成されます
a) 合皮 - 心皮はその側面とともに成長し、いくつかのリングが形成されます(チューリップ)。 果実の中に巣がいくつか作られています。
b) 傍果皮 - 心皮は端で一緒に成長し、1 つのリング (ケシ)、または中央の部屋を形成します。
c) lysicarpous - 心皮は端で一緒に成長し、1つの部屋または空洞を形成し、子房の底から柱が突き出ており、その上に胚珠があり、次に種子(クローブ)があります。
13.胚珠 - 1 つまたは 2 つの外皮に囲まれた核を持つ種子の茎 (ケーブル) からなる比較的複雑な構造。 種に応じて、胎盤は 1 つから多数の胚珠で成長します。 発生中の胚珠は最初は完全に核から構成されますが、すぐに 1 つまたは 2 つの外皮層 (外皮) が現れ、一端に小さな開口部である小門が現れます (図 6)。

米。 6. 胚珠と胚嚢の形成のスキーム。
1、2、3、4 – 核の発生、始胞子細胞の分離と減数分裂、3 つの大胞子の死。 5、6、7、8 – 雌の配偶体の大胞子(残り)からの発生、つまり胚嚢。
胚珠発生の初期段階では、単一の二倍体大胞子球が核内に現れます。 それは有糸分裂的に分裂して、通常は線状四分子に配置された 4 つの半数体大胞子を生成します。 これで大胞子形成が完了します。 通常、3 つの大胞子が破壊され、小門から最も遠い 4 つ目の大胞子が雌の配偶体に成長します。
機能的な大胞子はすぐに核を犠牲にして拡大し始め、その核は有糸分裂で 3 回分裂します。 3回目の有糸分裂の終わりには、8つの娘核が4つずつ2つのグループに分かれています - 巨大配偶体の小柱端の近くと、反対側のカラザール端にあります。 各グループの 1 つの核が 8 核細胞の中心に移動します。 それらは極性と呼ばれます。 小柱端の残りの 3 つの核は、1 つの卵と 2 つの協力細胞からなる卵装置を形成します。 カラザールの端でも、ここにある核の周囲に細胞膜が形成され、いわゆる対蹠細胞が発生します。 極核は二核の中央細胞に残ります。 この 8 核、7 細胞の構造は、胚嚢と呼ばれる成熟した女性の配偶体です。
5年生
装置:花のレイアウト。 黄色と緑色の粘土、アルミニウムと銅線、がく片と花びら用の紙ブランク。
5年生でこの教材を学習するときの目標は、生徒に花の構造を紹介し、「花被」と「花の主要部分」の概念を確実に理解させることです。 提案された実践的な部分は、観察と比較のスキルを開発し、正確さを養うための条件を作成します。
授業中
I. 組織化の瞬間
II. 宿題の確認
臓器とは何ですか? 臓器に名前を付けます 開花植物そしてその機能について説明します。
(p. 160、§40 の顕花植物の器官と機能に関する表への記入の正確さを確認します。)
Ⅲ. 参考知識の更新
顕花植物の名前はどのようにして付けられたのでしょうか? これらの植物の別名は何ですか? なぜ?
IV. 新しい教材の学習
先生は花の部分に名前を付け、その構造と機能について話します。 グループの生徒たちは説明を聞いた後、粘土で花の該当部分を作り、その名前をノートに書きます。 その結果、各グループは花のモデルを組み立てます。
ノートに書く

V. 統合
花の部分を列挙します。 モデル上でそれらを見つけてください。
花の主要な部分は何ですか? なぜ?
おしべとめしべは何でできていますか?
花被片とは何ですか? その機能は何ですか?
上記のようにノートに花をスケッチし、花の部分を表す番号を適切な順序で並べます。
VI. 宿題
対応する段落、メモ、ノートの図を調べてください。
6年生
装置:テキスト (付録を参照) – グループごとに 1 つ。 5年生で作った花の模型。 スライド – 透明フィルムまたは電子形式、およびスライドをデモンストレーションするための手段 (オーバーヘッド プロジェクター、ビデオ出力付きコンピューター、テレビまたはマルチメディア プロジェクター)。
6 年生でこの教材を学習するときの目標は、花についての知識を広げ、定着させ、花の分類に関する知識を確実に吸収することです。 仕事は、比較、分析し、結論を引き出す能力の発達に貢献します。 コミュニケーション文化、美的感覚、主題に対する認知的関心の発達を促進します。
授業中
I. 組織化の瞬間
クラスは5~6人のグループに分かれます。
II. 参考知識の更新
私たちは顕花植物の研究を続けています。 顕花植物と呼ばれる植物は何ですか? 顕花植物の別名は何ですか? なぜ? 開花植物の器官に名前を付けますが、まずそれらが 2 つのグループに分けられることを思い出してください。

これらの名前を説明してください。

どの臓器が栄養器官に分類されますか? 逃避とは何ですか?
フレーズを続けます: 器官は生成と呼ばれます...
Ⅲ. 新しい教材の学習
今日のレッスンの目標は、花についての既存の概念を拡張することです。
ノートブックのエントリ:"花"。

さまざまな植物の花はそれぞれサイズが異なります。目の前にあるのは最大の花であるラフレシア(直径約1メートル)で、最小の花であるウキクサ(植物全体がサムネイルに収まります)です。 花の形や色、部分の数、構造上の特徴が異なります。 しかし、類似点もあります。
付録の本文を読むと、すでによく知っている概念に遭遇するでしょう。 それらをもう一度繰り返します。 勉強する 新しい素材、次の質問について考えてください。
花は花被の構造によってどのようなグループに分けられますか?
花の主要部分の存在に基づいて、花はどのグループに分類できますか?
(学生のテキストを使った作品.)
V. 初期理解の確認

花の各部分に名前を付け、その構造と機能の特徴を簡単に説明します。
花被片とは何ですか?
花被片にはどんな種類があるの?
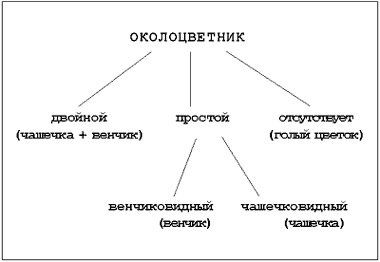
ノートブックのエントリ:花被片:二重(萼+花冠)。 シンプル(花冠形、カップ形)。 不在(裸の花)。
花のどの部分を主要部分と呼びますか? なぜ?
花は主要部分の存在に基づいてどのグループに分類されますか?

ノートブックのエントリ:花(主要部分の存在に基づく):両性(雄しべ+雌しべ)。 雌雄異体(雌雄異株)。 (めしべ); 無性(雄しべや雌しべがない)
雌雄同株と雌雄異株の植物とは何ですか?
V. 新しい知識の統合と応用
では、今日のレッスンで得た知識を実際に応用してみましょう。 花のモデルを使ってみましょう。
5年生で作った花の模型を見て、主要な部分の有無や果皮の構造から、目の前にある花は何なのかを判断してください。 ( 両性花被片を持つ.)
生徒の各グループは、元の花を修正する必要があるタスクが記載されたカードを受け取ります。
タスク:
1番目のグループ – 単純な花冠の形をした花被を持つ両性花。
2番目のグループ – シンプルなカップ型の花被を持つ両性花。
3番目のグループ – 二重花被を持つ雌花。
4番目のグループ – 二重花被を持つ雄花。
5番目のグループ – 女性(または男性)の裸の花。
グループ 6 – 無性花。
各グループは自分の花を展示し、モデルのどの部分を削除したのか、そしてその理由を説明します。 対応するスライドが同時に表示されます。
質問 ( グループ 3 とグループ 4 の花のモデルをデモンストレーションした後): カボチャの花は雌花と雄花のどちらが不毛の花と呼ばれますか? なぜ?
VI. 宿題
対応する段落を読んでください。
ノートに、癒合していないがく片 5 枚、癒合していない花弁 5 枚、雄しべ 5 つ、雌しべ 1 つを持つ通常の両性花の式を書き留めます。
「花」というテーマでクロスワード パズルを作成します (オプション)。
応用。
授業での課題に使用するテキスト
ドイツの偉大な詩人ゲーテでさえ、花を新芽が変化したものとみなすことを提案しました。 実際、花と新芽の構造には類似点があります。花柄と花托は花の茎の部分であり、萼、花冠、雄しべ、雌しべは葉が変形して形成されます。
したがって、花は種子(有性)繁殖に役立つ、短くなったシュートが改変されたものです。 他のシュートと同様に、花もつぼみから成長します。 通常、メインシュートまたはサイドシュートは花で終わります。
花の構造
レセプタクル - 花の軸。成長するにつれて、平ら、凹面、凸面など、さまざまな形を取ることができます。下の容器は、 花柄 - ほとんどの植物で花が着く細い茎。 多くの植物の花柄には、2枚(双子葉植物の場合)または1枚(単子葉植物の場合)の小さな葉が発達します。これは、 托葉。 花柄のない花を無柄花といいます。
花被片 で構成されます。 萼 そして カローラ 呼ばれた ダブル。 これは、サクラ、キャベツ、バラ、その他多くの植物の花の花被です。

カップ
花被の外円を形成します。 萼は通常、小さな緑色の葉で構成されています。 がく片(H)。 カーネーションなどの一部の植物では、がく片の下部が一緒になって筒状に成長します。これは萼と呼ばれます。 プレキシフォリア。 他のもの、例えばゼラニウムではがく片が一緒に成長しません。 ディオフィロスカップ。
花が咲くとガクが落ちる場合もありますが、開花中はガクが残っていることが多いです。
カップは次の機能を実行します。
1)つぼみが開くまで花の内部を保護します。
2) 光合成のプロセスは緑色のがく片で起こります。
泡立てます
- 花被の内側の部分は、通常、明るい色の大きな花序で構成されています。 花びら(L)。 一部の植物(香りのよいタバコ、ナス科の植物、サクラソウ)では、花びらが一緒に成長して形成されます。 花弁の間の泡立てます。 他のもの(キャベツ、リンゴ、サクランボ)では、花冠は別々の花びらで構成されており、 別々の花びらを持つ、 または 自由な花びらのある.
いくつかの植物、主に単子葉植物 (ユリ、アマリリス、チューリップ) では、すべての花被片は多かれ少なかれ同じです。 この花被片はと呼ばれます 単純(について)。 たとえば、チューリップ、ハシバミライチョウ、蘭などの一部の植物では、単純な花被の花被片は花びらのように大きくて明るいです。 単純な花冠の形をした花被片.

他の植物、たとえばイグサ、ビート、イラクサでは、単純な花被の葉は小さく、目立たず、通常は緑色で、がく片に似ています。 シンプルなカップ状の花被片.
泡立て器の主な機能:
1) 受粉昆虫を誘引する。
2) 花の主要部分の保護。
花被のない花(トネリコ、スゲ、ヤナギ)もあり、それらは呼ばれます 裸.
花被片(一重および二重)は、それを通していくつかの対称軸を描くことができるように配置できます(リンゴの木、桜の木、キャベツなど)。 このような花はこう呼ばれます 正しい。 1 つの対称軸を描くことができる花 (エンドウ豆、セージ)、または対称軸を描かない花 (カンナ) を呼びます。 間違っている.
乳棒 (P)と 雄しべ (た) - 花の主要な部分、生殖細胞-配偶子-がその中で形成されます。
おしべ からなる フィラメント、それの助けを借りて、それは容器に付着し、雄の配偶子を持つ花粉を含む葯 - 精子。 花糸がなく、葯が容器に直接位置する場合、それはと呼ばれます 座りがちな.

乳棒 に分け 汚名(上部は特殊な生地で作られており、花粉をキャッチします)、 カラムそして 卵巣(雌の配偶子である卵が成熟する下部の拡張部分)。 雌しべに花柱がなく、柱頭が子房にある場合は、と呼ばれます。 座りがちな。 花のめしべから種のある果実ができます。

ほとんどの植物には、雄しべと雌しべの両方を含む花があります。 これ 両性花。 しかし、一部の植物(キュウリ、トウモロコシ)では、雌しべしかない花もあります。 めしべ、 または 婦人向け, フラワーズ、その他はおしべだけですが、これらは スタミナテ、 または スタミナテ、, フラワーズ。 このような花はこう呼ばれます 雌雄異株.
無性愛者これらは、雄しべと雌しべの両方の主要な部分をすべて欠いている花と呼ばれます。 それらは、主要な部分を持つ他の花に受粉昆虫を誘引する機能のみを実行します。 無性花はヤグルマギク、ヒマワリ、その他の植物の花序に見られます。
いくつかの花には 蜜腺- 甘い液体を生成する腺。 ほとんどの場合、それらはレセプタクル上にあります。
花の構造を象徴するために数式が使用されます。 式をコンパイルするには、次の表記を使用します。
O – 単純な花被片。
Ch – がく片。
L – 花びら:
T – 雄しべ。
P – 乳棒。
がく片、花びら、おしべ、めしべの数を数字で示し、12個以上の場合は記号で示します。
花の一部が一緒に成長している場合は、対応する番号が括弧内に書き込まれます。
正しい花はアスタリスク * で示されています。
不正確 - 矢印;
単性の雄(雄蕊)花 - ;
女性(雌しべ) – サイン。
バイセクシュアル - サイン。
花は顕花植物の中で目立つ、多くの場合美しく、重要な部分です。 花は大きい場合もあれば小さい場合もあり、明るい色や緑色、香りがあるか無臭である場合もあり、単独で咲いている場合もあれば、多くの小さな花が集まって 1 つの共通の花序になっている場合もあります。
花は、種子の繁殖に使用される、短くなったシュートが改変されたものです。 通常、メインシュートまたはサイドシュートは花で終わります。 他のシュートと同様に、花もつぼみから成長します。
花の構造
花 - 生殖器官 被子植物短くなった茎(花軸)からなり、その上に花被(花被)、雄しべ、雌しべがあり、1つ以上の心皮で構成されています。
花の軸のことを「花軸」といいます。 レセプタクル。 受け皿は成長し、 違う形平ら、凹面、凸面、半球形、円錐形、細長い、円柱形。 下の容器は花柄に変わり、花と茎または花柄を接続します。
花柄のない花を無柄花といいます。 多くの植物の花柄には、2つまたは1つの小さな葉、つまり苞があります。
花の表紙 - 花被片- 萼と花冠に分けることができます。
カップ花被の外側の円を形成し、その葉は通常比較的小さいサイズです。 緑色。 萼は分離したものと融合したものがあります。 通常、つぼみが開くまで花の内部を保護する役割を果たします。 場合によっては、花が開くときにガクが落ちますが、ほとんどの場合、開花中に残ります。
花の雄しべと雌しべの周りにある部分を花被といいます。
内側の小葉は花冠を構成する花びらです。 外側の葉 - がく片 - は萼を形成します。 花被片は萼と花冠からなり、八重と呼ばれます。 花冠とがくに分かれていない花被で、花の小葉はすべてほぼ同じで、単純です。

泡立てます- 花被の内側の部分で、萼とは明るい色と大きなサイズが異なります。 花びらの色は色素体の存在によるものです。 花冠には分離した花冠と融合した花冠があります。 最初のものは個々の花びらで構成されます。 融合した花弁の花冠では、管が区別され、それに垂直に位置する手足があり、一定の数の歯または花冠の刃があります。
花は対称または非対称にすることができます。 花被のない花もあり、裸花と呼ばれます。
対称(アクノモーフィック)- リムを通して多くの対称軸を引くことができるかどうか。
非対称(接合形)- 対称軸が 1 つだけ描画できる場合。
八重咲きの花びらの枚数が異常に増えています。 ほとんどの場合、花びらが分裂した結果として発生します。
おしべ- 花の一部で、小胞子や花粉を形成する特殊な構造の一種です。 それは、それを介して容器に付着するフィラメントと、花粉を含む葯で構成されます。 花のおしべの数は体系的な特徴です。 雄しべは、容器への付着方法、形状、サイズ、雄しべのフィラメントの構造、結合組織、および葯によって区別されます。 花の中の雄しべの集まりはアンドロエシウムと呼ばれます。
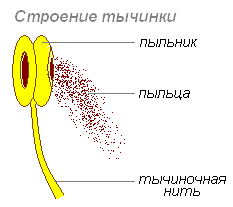
フィラメント- 雄しべの無菌部分で、頂点に葯があります。 フィラメントは、真っ直ぐ、曲がっている、ねじれている、曲がりくねっている、または壊れている場合があります。 形状:髪の毛状、円錐形、円筒形、平ら、棍棒形。 表面の性質は、裸で、思春期で、毛深く、腺があります。 植物によっては、それが短いか、まったく発育しない場合もあります。
葯フィラメントの上部に位置し、結合組織によってフィラメントに付着しています。 コネクタによって相互に接続された 2 つの部分で構成されます。 葯の各半分には、花粉が発達する 2 つの空洞 (花粉嚢、部屋、または巣) があります。

葯は原則として4室ですが、場合によっては各半分の巣の間の隔壁が壊れて2室になることがあります。 植物によっては、葯が単葉である場合もあります。 巣が3つある場合は非常にまれです。 花糸への付着の種類に基づいて、葯は固定葯、可動葯、および振動葯に分類されます。
葯には花粉または花粉粒が含まれています。
花粉粒子の構造
雄しべの葯で形成される塵の粒子は花粉粒と呼ばれます。 最大のものは直径0.5 mmに達しますが、通常ははるかに小さいです。 顕微鏡で観察すると、さまざまな植物から採取された塵の粒子がまったく同じではないことがわかります。 大きさや形が異なります。

塵粒子の表面はさまざまな突起や結節で覆われています。 花粉は雌しべの柱頭に付着すると、伸長物と柱頭に分泌される粘着性の液体の助けを借りて保持されます。
若い葯の巣には特別な二倍体細胞が含まれています。 減数分裂の結果、各細胞から 4 つの一倍体胞子が形成されます。これらは、サイズが非常に小さいため小胞子と呼ばれます。 ここで、花粉嚢の空洞内で、小胞子が花粉粒に変わります。

これは次のように起こります。小胞子核は有糸分裂により栄養核と生殖核の 2 つに分割されます。 細胞質の領域は核の周囲に集中し、栄養細胞と生殖細胞の2つの細胞が形成されます。 小胞子の細胞膜の表面には、酸やアルカリに不溶な非常に強力な殻が花粉嚢の内容物から形成されます。 したがって、各花粉粒は栄養細胞と生殖細胞で構成され、2 つの膜で覆われています。 植物の花粉は多くの花粉粒で構成されています。 花粉は花が開くと同時に葯の中で成熟します。
花粉の発芽
花粉の発芽の開始は有糸分裂に関連しており、その結果、小さな生殖細胞(そこから精子細胞が発達)と大きな栄養細胞(そこから花粉管が発達)が形成されます。
花粉が何らかの方法で柱頭に到達すると、発芽が始まります。 柱頭の粘着性と凹凸のある表面は花粉を保持するのに役立ちます。 さらに、柱頭は花粉に作用する特殊な物質(酵素)を分泌し、花粉の発芽を促します。
花粉が膨張し、外皮(花粉粒の殻の外層)の抑制的な影響により、花粉細胞の内容物が孔の1つを破り、その孔を通って内膜(花粉粒の内側の孔のない殻)が開きます。細い花粉管の形で外側に突き出ます。 花粉細胞の内容物は花粉管に入ります。
柱頭の表皮の下には緩い組織があり、そこに花粉管が侵入します。 それは、粘液細胞間の特別な伝導チャネルを通過するか、柱の伝導組織の細胞間空間に沿って曲がりくねって成長し続けます。 この場合、通常、かなりの数の花粉管が花柱内を同時に前進し、いずれかの花粉管の「成功」は個々の成長速度に依存します。
2つの精子と1つの栄養核が花粉管の中を通過します。 花粉内で精細胞の形成がまだ起こっていない場合、生殖細胞が花粉管に入り、ここでその分裂によって精細胞が形成されます。 多くの場合、栄養核は管の成長端の前方に位置し、精子はその後ろに連続して位置します。 花粉管の中では、細胞質が絶えず運動しています。

花粉には栄養が豊富に含まれています。 これらの物質、特に炭水化物(砂糖、デンプン、ペントサン)は、花粉の発芽中に集中的に消費されます。 炭水化物に加えて、 化学組成花粉には、タンパク質、脂肪、灰分、および多数の酵素が含まれています。 花粉には含まれています 高いコンテンツリン。 花粉中の物質は移動可能な状態にあります。 花粉が移りやすい 低温最大-20℃、さらにはそれ以下の環境で長時間使用できます。 高温発芽をすぐに減らします。
乳棒
雌しべは花の果実を形成する部分です。 それは心皮(胚珠を有する葉のような構造)から生じ、その後後者の端が融合します。 1 つの心皮で構成されている場合は単純ですが、側壁と融合した複数の単純な雌しべで構成されている場合は複雑になります。 一部の植物では、雌しべが未発達で、原始的なものだけで表されます。 雌しべは子房、花柱、柱頭に分かれます。


卵巣 — 下部雌しべ、種子の芽が入っている部分。
子房に入った花粉管はさらに成長し、ほとんどの場合、花粉管(小丘)を通って胚珠に入ります。 胚のうに侵入すると、花粉管の端が破裂し、内容物が協力剤の1つにこぼれ、それが黒くなり、すぐに崩壊します。 栄養核は通常、花粉管が胚のうに侵入する前に破壊されます。
花は定期的と不規則的
花被片(単純および二重)は、それを通していくつかの対称面を描くことができるように配置できます。 このような花はレギュラーと呼ばれます。 1 つの対称面を描くことができる花を不規則と呼びます。
花は両性で雌雄異株です
ほとんどの植物には、雄しべと雌しべの両方を含む花があります。 これ 両性花。 しかし、植物によっては、雌しべだけを持つ花(雌蕊花)もあれば、雄しべだけを持つ花(雄蕊花)もあります。 このような花は雌雄異株と呼ばれます。
雌雄異株と雌雄異株の植物
雌蕊花と雌蕊花の両方を咲かせる植物を雌雄同株と呼びます。 雌雄異株の植物は、ある植物には雌蕊の花があり、別の植物には雌蕊の花があります。
同じ植物に両性花と単性花が見られる種があります。 これらはいわゆる一夫多妻(一夫多妻)植物です。
花序
花は新芽に形成されます。 単独で存在することは非常にまれです。 はるかに多くの場合、花は花序と呼ばれる目立つグループに集められます。 花序の研究はリンネから始まりました。 しかし彼にとって、花序は一種の枝分かれではなく、開花の方法でした。
花序は主軸と横軸(無柄または花柄)で区別され、そのような花序は単純と呼ばれます。 花が横軸上にある場合、これらは複雑な花序です。
| 花序の種類 | 花序図 | 特徴 | 例 |
| 単純な花序 | |||
| みがきます | 個々の側花は細長い主軸の上にあり、同時に長さがほぼ等しい独自の花柄を持ちます。 | バードチェリー、スズラン、キャベツ | |
| 耳 | 主軸は多かれ少なかれ細長くなりますが、花には茎がありません。 固着性の。 | オオバコ、蘭 | |
| 穂軸 | 耳と異なるのは、軸が太くて肉質であることです。 | とうもろこし、書道 | |
| バスケット | 花は常に固着性で、短くなった軸の強く肥厚して広がった端に位置し、凹面、平坦または凸面の外観を持ちます。 この場合、外側の花序にはいわゆる総苞があり、遊離または融合した 1 列または複数の連続する苞葉から構成されます。 | カモミール、タンポポ、アスター、ヒマワリ、ヤグルマギク | |
| 頭 | 主軸は大幅に短くなり、側花は固着性またはほぼ固着性で、互いに狭い間隔で配置されます。 | クローバー、スカビオサ | |
| 傘 | 主軸が短くなります。 側花はあたかも一か所から出現し、同じ平面またはドーム形に位置する異なる長さの茎の上に座ります。 | サクラソウ、タマネギ、チェリー | |
| シールド | 総状花序との違いは、下の花に長い花柄があり、その結果、花がほぼ同一平面上に位置することです。 | 梨、シモツケ | |
| 複雑な花序 | |||
| 複雑なブラシまたは泡立て器 |  | 側枝軸は主軸から伸びており、その上に花または単純な花序が配置されています。 | ライラック、オーツ麦 |
| 複雑な傘 | 短くなった主軸から単純な花序が伸びます。 | にんじん、パセリ | |
| 複雑な耳 | 個々の小穂は主軸上にあります。 | ライ麦、小麦、大麦、ウィートグラス | |
花序の生物学的意義
花序の生物学的重要性は、小さく目立たない花が集められると目立つようになり、大量の花粉を生成し、花から花へと花粉を運ぶ昆虫をよりよく引き付けることです。
受粉
受精が起こるためには、花粉が柱頭に着地する必要があります。
花粉を雄しべから雌しべの柱頭に移すプロセスを受粉といいます。 受粉には、自家受粉と他家受粉の 2 つの主なタイプがあります。
自家受粉
自家受粉では、雄しべからの花粉が同じ花の柱頭に付着します。 これが小麦、米、オート麦、大麦、エンドウ豆、豆、綿花の受粉方法です。 植物の自家受粉は、ほとんどの場合、まだ開いていない花、つまり花が開いたときにはすでに終わっているつぼみの中で行われます。
自家受粉中に、同じ植物上で形成された性細胞が形成されるため、同じ遺伝的特徴を持つ性細胞が融合します。 これが、自家受粉のプロセスから生じる子孫が親植物と非常に似ている理由です。

他家受粉
他家受粉中に、父方生物と母方生物の遺伝的特徴の組み換えが起こり、その結果として生じる子孫は、親が持っていなかった新しい特性を獲得することがあります。 そのような子孫はより生存可能です。 自然界では、他家受粉は自家受粉よりもはるかに頻繁に発生します。

他家受粉は、さまざまな外部要因の助けを借りて行われます。
風友病(風による受粉)。 風媒性植物では、花は小さく、花序に集まることが多く、花粉が多量に生産され、乾燥していて小さく、葯が開くと勢いよく吐き出されます。 これらの植物からの軽い花粉は、風に乗って最大数百キロメートルの距離まで運ばれる可能性があります。

葯は長くて細い糸状体の上にあります。 雌しべの柱頭は幅広または長く、羽毛状で花から突き出ています。 風気症は、ほとんどすべての草とスゲの特徴です。
昆虫好き(昆虫による花粉の移送)。 植物の昆虫食性への適応は、花の匂い、色、大きさ、伸びた粘着性の花粉などです。 ほとんどの花は両性花ですが、花粉と雌しべの成熟が同時に起こらなかったり、柱頭の高さが葯の高さよりも高かったり低かったりして、自家受粉を防ぐ役割を果たしています。
昆虫によって受粉される植物の花には、甘くて芳香のある溶液を分泌する領域があります。 これらの領域は蜜腺と呼ばれます。 蜜腺は花のさまざまな場所にあり、 さまざまな形。 花に飛来した昆虫は蜜腺や葯に引き寄せられ、食事中に花粉で汚れます。 昆虫が別の花に移動すると、昆虫が運ぶ花粉が柱頭に付着します。
昆虫によって受粉すると、無駄にされる花粉が少なくなるため、植物は花粉の生産量を減らして栄養素を節約します。 花粉は空気中に長時間滞留する必要がないため、重くなる可能性があります。
昆虫は、まばらに配置された花や風のない場所、つまり森の茂みの中や厚い草の中の花に受粉することができます。
通常、各植物種は数種類の昆虫によって受粉され、各種類の受粉昆虫は複数の植物種に役立ちます。 しかし、1 種類の昆虫のみによって花が受粉される植物もあります。 このような場合、花と昆虫の生態や構造の相互対応は、奇跡のように思えるほど完璧です。
鳥類愛好症(鳥による受粉)。 一部の人にとっては典型的 熱帯植物色鮮やかな花、豊富な蜜分泌、強い弾力性のある構造。
親水性(水による受粉)。 で観察されました 水生植物。 これらの植物の花粉と柱頭は、ほとんどの場合糸状の形状をしています。
獣姦(動物による受粉)。 これらの植物は、花の大きさ、粘液を含む蜜の豊富な分泌、受粉時の花粉の大量生産によって特徴付けられます。 コウモリ- 夜に咲きます。
受精
花粉粒は雌しべの柱頭に着地し、殻の構造的特徴と、花粉が付着する柱頭の粘着性の甘い分泌物によって雌しべに付着します。 花粉粒は膨張して発芽し、長くて非常に細い花粉管になります。 花粉管は、栄養細胞の分裂の結果として形成されます。 まず、この管は柱頭の細胞間で成長し、次に花柱に成長し、最後に卵巣の腔内に成長します。
花粉粒の生殖細胞が花粉管内に移動し、分裂して2つの細胞を形成します。 雄の配偶子(精子)。 花粉管が花粉管を通って胚のうに侵入すると、精子の1つが卵子と融合します。 受精が起こり、接合子が形成されます。
2 番目の精子は、胚嚢の大きな中央細胞によって核と融合します。 したがって、顕花植物では、受精中に 2 つの融合が起こります。最初の精子は卵子と融合し、2 番目の精子は大きな中央細胞と融合します。 このプロセスは 1898 年にロシアの植物学者で学者の S.G. ナヴァシンによって発見され、それと呼ばれました。 重複受精。 重複受精は顕花植物のみの特徴です。
配偶子の融合によって形成された接合子は 2 つの細胞に分かれます。 生じた各細胞は再び分裂するなどして、細胞分裂が繰り返された結果、新しい植物の多細胞胚が発生します。
中央細胞も分裂して、栄養貯蔵が蓄積する胚乳細胞を形成します。 それらは胚の栄養と発育に必要です。 種皮は胚珠の外皮から発達します。 受精後、種子は胚珠から発生し、皮、胚、栄養源から構成されます。
受精後は卵巣に流れます 栄養素、徐々に熟した果実になっていきます。 種子を守る果皮 有害な影響、卵巣の壁から発生します。 一部の植物では、花の他の部分も果実の形成に関与します。

教育紛争
雄しべでの花粉の形成と同時に、胚珠で大きな二倍体細胞の形成が起こります。 この細胞は減数分裂で分裂し、小胞子よりもサイズが大きいため大胞子と呼ばれる 4 つの一倍体胞子を生じます。

形成された 4 つの大胞子のうち 3 つが死に、4 つ目は成長し始め、徐々に胚嚢に変化します。
胚嚢の形成
核の 3 回の有糸分裂の結果、胚嚢の空洞内に 8 つの核が形成され、細胞質で覆われます。 膜を失った細胞が形成され、特定の順序で配置されます。 胚嚢の一方の極では、卵と 2 つの補助細胞からなる卵装置が形成されます。 反対極には 3 つのセル (対蹠体) があります。 1 つの核が各極から胚嚢の中心に移動します (極核)。 場合によっては、極核が融合して胚嚢の二倍体中心核を形成します。 核分化が起こった胚嚢は成熟したとみなされ、精子を受け入れることができます。

花粉と胚嚢が成熟するまでに花が開きます。
胚珠の構造
胚珠は成長します 内側の側面子房の壁は、植物の他の部分と同様に細胞で構成されています。 植物によって子房内の胚珠の数は異なります。 小麦、大麦、ライ麦、サクランボでは、子房には胚珠が1つしか含まれていませんが、綿では数十個、ケシではその数は数千に達します。
それぞれの胚珠はカバーで覆われています。 胚珠の上部には花粉の通り道である細い管があります。 それは胚珠の中心部分を占める組織につながります。 この組織では、細胞分裂の結果として胚嚢が形成されます。 花粉開口部の反対側には卵細胞があり、中央部分は大きな中央細胞で占められています。
被子植物(開花)植物の発達

種子と果実の形成
種子と果実が形成されると、精子の 1 つが卵子と融合し、二倍体接合子を形成します。 その後、受精卵は何度も分裂し、その結果、多細胞植物の胚が発生します。 2番目の精子と融合した中央細胞も何度も分裂しますが、2番目の胚は発生しません。 特別な組織、つまり胚乳が形成されます。 胚乳細胞は、胚の発育に必要な栄養素を蓄積します。 胚珠の外皮が成長して種皮になります。

したがって、重複受精の結果として、胚、貯蔵組織(胚乳)および種皮からなる種子が形成されます。 子房の壁は果皮と呼ばれる果実の壁を形成します。
有性生殖
被子植物の有性生殖は花と関連しています。 その最も重要な部分は雄しべと雌しべです。 それらは起こります 複雑なプロセス有性生殖に関係します。
開花植物では、雄の配偶子 (精子) は非常に小さいのに対し、雌の配偶子 (卵) ははるかに大きくなります。
おしべの葯では細胞分裂が起こり、花粉粒が形成されます。 被子植物の各花粉粒は、栄養細胞と生殖細胞で構成されています。 花粉粒は2回のコートで覆われています。 通常、外殻は不均一で、棘、いぼ、網目状の突起があります。 これは、花粉粒が柱頭に留まるのに役立ちます。 植物の花粉は、葯の中で成熟し、花が開くまでに多くの花粉粒で構成されます。
フラワーフォーミュラ
数式は、花の構造を条件付きで表現するために使用されます。 花の式をコンパイルするには、次の表記法を使用します。
がく片のみ、または花弁のみからなる単純な花被片で、その一部を花被片と呼びます。 |
|
| H | 萼、萼片からなる |
| L | 花冠は花びらで構成されています |
| T | おしべ |
| P | 乳棒 |
| 1,2,3... | 花の要素の数を数字で示します |
| , | 花の同じ部分は形が異なります |
| () | 花の融合部分 |
| + | 2 つの円内の要素の配置 |
| _ | 上部または下部の子房 - 雌しべの数を示す数字の上または下の線 |
| 間違った花 | |
| * | 適切な花 |
| ♂ | ユニセックスのスタミナテフラワー |
| ♀ | 雌雄異株の花 |
| バイセクシャル | |
| ∞ | 花の部分の数が12を超える |
桜の公式の例:
*H 5 L 5 T ∞ P 1
花図
花の構造は数式だけでなく図でも表すことができます。 模式図花を花の軸に垂直な平面上に配置します。

開いていない花芽の断面図を使って図を作成します。 この図は、式では示すことができない各部分の相対的な位置も示すため、式よりも花の構造についてより完全なアイデアを提供します。
花は複雑な器官システムであり、 種子の繁殖開花植物の中で。 進化の過程における花の出現はアロモルフォシスであり、これが原因となった 幅広い用途地球上の被子植物、または顕花植物。
花の役割:
- 胚珠を伴う心皮(雌しべ)の花粉粒を伴う雄しべの形成。
- 受粉。
- 複雑な受精プロセス。
- 種子と果実の形成。
花- これは、花被、雄しべ、および心皮(雌しべ)を持つ、成長が制限された短縮された改変されたシュートです。 すべての顕花植物の花の構造は似ており、その形はさまざまです。 ここに特異的適応、つまりへの適応が現れます。 さまざまな方法で受粉。
花の外部構造
花は主茎または側茎で終わります。 花の下の茎の葉のない部分は、と呼ばれます。 花柄。 固着花では花柄がないか、または著しく短くなります。 花柄は花の短くなった軸、その茎の部分に入ります。 レセプタクル。 レセプタクルの形状は、細長、凸状、平坦、凹状とすることができます。 花びらには、がく片と花びら、おしべとめしべなど、花のすべての部分が含まれています。
萼片と花弁が一緒になって構成されます 花被片。 がく片は通常、花、特につぼみを損傷から保護しますが、他の機能も果たします。 光合成は葉緑体を含む緑色のがく片で起こります。 一部の植物(チューリップ、アネモネ)では、それらは花びらの形になり、花びらの機能を果たします。 発育中の果物を保護し、その流通を保護するのに役立ちます。
がく片は上部の栄養葉から始まります。 その証拠は、葉との形態学的類似性であり、一部の植物 (牡丹) で明確に表現されており、それらの螺旋状の配置が挙げられます。 がく片が組み合わさって萼を形成し、雌雄異株または融合葉になります。

花びら花粉媒介者を引き付け、受粉の成功を促進する機能を果たします。 花びらの起源は 2 つあり、植物によっては雄しべが変化したものもあります。 このような花びらは、スイレンのほか、キンポウゲ科、ナデシコ科、ケシ科などの代表的な植物に見られます。別のグループの植物には、葉由来のがく片のような花弁があります(牡丹、マグノリア)。
花びらの集まりを「花びら」といいます。 泡立てます。 花冠の大きさ、構造、色は多様であり、これは受粉の生物学に関連しています。 風によって受粉する植物では、花冠は未発達であるか、存在しません。 花びらは端で一緒に成長し、速く花びらを広げる花冠 (ヒルガオ、ペチュニア) を形成することがあります。 進化の過程で、このような花冠は、花弁のない花冠から進化しました。
花に萼と花冠がある場合、花被片は八重と呼ばれます。 花びらがない場合、または花びらの違いが明確に表現されていない場合、花被は単純と呼ばれます。 シンプルな花被は、チューリップ、ユリ、スズランのような明るい色の花冠の形、または麻、キノア、イラクサのカップ形の緑色のいずれかです。 花被のない花は裸と呼ばれます - スゲ、ヤナギ。

花被の内側の花びらに近い部分にあります。 雄しべ。 その数は、1 つから 10 つ以上までさまざまです。 進化の過程で、雄しべは花糸と葯に分化しました。 葯は、フィラメントの続きによって接続された 2 つの半分から構成されます。 葯の各半分には 2 つの胞子嚢が含まれており、それらは葯巣、または花粉嚢と呼ばれます。

巣は一次胞子形成細胞からの組織で満たされています。 一連の連続的な有糸分裂の結果、一次胞子形成細胞から多くの母細胞、つまり小胞子が形成されます。 その後、母細胞は減数分裂的に分裂し、四分体の一倍体小胞子を形成します。 このような各小胞子は花粉粒に変わります。 これを行うために、サイズが大きくなり、外側(外皮)と内側(内膜)の二重の殻で覆われます。 外殻は、その主成分であるスポロポレニンのおかげで、高い耐性を特徴としています。酸やアルカリに溶けず、300℃までの温度に耐えることができ、地層の堆積物で何百万年も保存されます。
雄性配偶体は花粉粒の内部で形成されます。一倍体小胞子は有糸分裂的に分裂し、より大きな管細胞 (栄養細胞) とその中に小さな生殖細胞を形成します。 生殖細胞は再び有糸分裂的に2つの雄の配偶子、つまり精子に分裂します。

花の内部は占有されています めしべ。 その数は 1 つから 10 つ以上までさまざまです。 各雌しべは、1 つまたは多数の融合した心皮によって形成されます。
雌しべの一番下、つまり子房には胚珠(胚珠)があります。 進化の過程で、その上部から柱が形成され、雌しべの上に柱頭が上がります。 スタイルがない場合、その汚名は固着と呼ばれます。 子房が平らな容器または凸状の容器の上に位置し、花の他のすべての部分が雌しべの下に取り付けられている場合、子房が優れている可能性があります。 下位の子房を持つ花では、凹面の容器がその壁と融合し、花被と雄しべが雌しべの上に取り付けられます。

雌しべの子房には空洞、つまり巣があります。 卵巣には単房性と多房性があります。 多房性の子房は、いくつかの心皮の融合の結果として形成されます。 巣の数は、融合した心皮の数に等しい。 各巣では、胚珠(胚珠)が子房の壁に固着または柄の上に形成されます。 1本(梅、桜)から数千本(ケシ、蘭)まであります。
胚珠(胚珠)の構造
胚珠の解剖学的検査により、次の構成要素が区別されます。
- ケーブルカー;
- 核;
- カバー。
- ミクロパイル;
- 胎芽嚢。
による 種子の茎栄養分が胚嚢に入り、胚珠が卵巣に付着します。 核核胚珠は、大胞子に栄養を与え、保護する実質組織です。 外側では、核は1つまたは2つで覆われています カバー(外皮)。 核を完全に覆うわけではありません。 多くの場合、それらは胚珠の上で結合せず、胚珠と呼ばれる小さな穴を形成します。 ミクロパイル、または花粉の通過。
胚珠の最も内側の部分を占めます 胎芽嚢、被子植物では雌の配偶体です。

胚珠(胚珠)は、大胞子嚢と周囲の被膜で構成されています。 大胞子嚢では、1 つの母細胞が形成され、そこから減数分裂によって 4 つ組の半数体大胞子が形成されます。 そのうちの 3 つが死んで破壊され、4 つ目(雌の配偶体を生じさせる)の大胞子は長さが大幅に伸び、同時にその半数体核が有糸分裂によって分裂します。 娘核は、細長い細胞の異なる極に分岐します。
さらに、得られた各核は有糸分裂でさらに 2 回分裂し、細胞の異なる極に 4 つの半数体核を形成します。 これはすでに 8 つの一倍体核を持つ胚嚢です。 次に、2 つの四倍体核のそれぞれから 1 つが胚のうの中心に送られ、そこで融合して二次二倍体核が形成されます。
この後、胚嚢の細胞質に核間の細胞隔壁が現れ、7細胞になります。
胚のうの極の 1 つに、より大きな卵と 2 つの補助細胞からなる卵装置があります。 反対側の極には 3 つの対蹠細胞があります。 6 つの細胞はすべて一倍体です。 中心には二次核を持つ二倍体細胞があります。
ほとんどの植物では、花には雄しべと雌しべがあり、両性花と呼ばれます。 花も単性花で、雄花(雄花)または雌花(雌花)です。 雄花と雌花が1つの個体に存在する場合があり、そのような植物は雌雄異株(キュウリ、トウモロコシ、オーク、シラカバ)と呼ばれ、別の個体にある場合は雌雄異株(麻、ヤナギ、ポプラ)と呼ばれます。 単性花や雌雄異株の植物は他家受粉への適応の 1 つです。
プラント図とフォーラム
のために 簡単な説明花は図と公式を使用します。 図は、花の要素をその軸に垂直な平面に概略的に投影したものです。 花のすべての部分、苞および母芽は、特定の記号で指定されます。 がく片 - 巻き括弧付き、花弁 - 丸い括弧付き、雄しべ - 葯を通る横断面を持つ、雌しべ - 横断面を持つ卵巣を通って。
花の公式を作るときは、花被をO、がく片をH、花びらをL、雄しべをT、雌しべをPと表記します。花の部分の数は、根元に書かれた数字で示されます。手紙の。 おしべとめしべの数が12本以上の場合は、アイコン-∞を付けます。 花の一部が一緒に成長する場合、対応する番号が括弧内に入れられます。 上の子房は番号の下の水平線で示され、下の子房は雌しべの数の上にあります。







